
|

|



|
|
|
|
|
|
|
|
Handbuch der Zoologie/Handbook of Zoology.
Band/Volume IV Arthropoda: Insecta.
Teilband/Part 38 Coleoptera, Beetles.
Volume 1: Morphology and Systematics
(Archostemata, Adephaga, Myxophaga, Polyphaga partim).
R.G. Beutel & R.A.B. Leschen (eds.).
Walter de Gruyter: Berlin- N.Y., 1-567
Благодаря М.Г. Волковичу редакция получила возможность познакомиться с томом учебной серии по зоологии, в котором дан обзор последней литературы по общим вопросам колеоптерологии, и который включает несколько разделов по семействам жуков, причем каждое из попавших в этот том семейств сопровождается не только обзором литературы, но достаточно полной информацией о составе и объеме семейства, распространении, биономии, экологии, строении всех фаз жизненного цикла, филогении и таксономии. Разделы по многим семействам подготовлены ведущими специалистами, работающими с ними. Всего в подготовке тома участвовало 38 авторов, в том числе двое отечественных: В.В. Гребенников, работающий в настоящее время в Канаде, и М.Г. Волкович из ЗИНа. Многие общие разделы подготовлены редакторами тома или при их соавторстве (R.G. Beutel и R.A.B. Leschen) и поэтому позволяют судить о взглядах редакторов на многие аспекты колеоптерологии и науки в целом. Рассмотрение всех разделов книги требует много времени, и поэтому автор обзора нашел возможным ограничиться здесь только некоторыми важными моментами в содержании обильной информации книги, не пытаясь уделить равное внимание всем группам и всем разделам. Следует заметить, что многие семейства, попавшие в том, имеют скорее всего более обоснованный объем в сравнении с другими интерпретациями рангов и таксономического состава групп (они большей частью позаимствованы из сводки Lawrence & Newton, 1995). В частности, по-видимому, вполне оправдана трактовка семейства водолюбов (Hydrophilidae), согласно которой группы Helophorinae, Epimetopinae, Georissinae, Hydrochinae, Spercheinae включаются в него в ранге подсемейств, а не обособленных семейств (как это предпочитают делать другие специалисты). Также, по-видимому, оправдано широкое понимание семейства лейодид (Leiodidae), объединяющего также подсемейства Chelovinae, Platypsillinae и т.д. Вместе с тем, иерархическая композиция и соотношение рангов скарабеоидных (Scarabaeoidea), некоторых надсемейств элатериформных жуков (Elateriformia), а также таксономический состав и ранги внутри надсемейств Cucujoidea и Tenebrionoidea еще нуждаются в существенном совершенствовании. Редакторы в основном следуют компилятивным работам Дж.Ф. Лоуренса, даже в тех случаях, когда это не является очевидно оправданным. Принимается, например, положение Agnathus decoratus (Germar, 1818), которое ему дается в работе Lawrence & Newton, 1995, но не другие не менее резонные интерпретации (см. обзор на нашем сайте "К какому же семейству относится Agnathus decoratus?"). Хотя редакторы утверждают, что они включили в их список семейств группы, введенные после Lawrence et al., 1999, ряд досадных пробелов в нем все же обнаруживается. Можно, например, допустить, что редактора не приняли предложение С.В. Казанцева поднять статус группы рода Dexoris Waterhouse, 1878 до уровня семейства Dexoridae Kleine, 1933 (Kazantsev, 2003), но непонятно - почему в списке отсутствует подсемейство Maynipeplinae Kirejtshuk, 1998, которое есть в том числе и в работе Lawrence et al., 1999 ?. Таким образом, неизвестно, к названию какого таксона редакторы синонимизировали последнее из упомянутых названий. Представляется, что было бы оправданным включение в предложенный редакторами список всех синонимов для семейств и подсемейств, которые облегчили бы читателям восприятие их систематической интерпретации.
Небольшой, но достаточно содержательный раздел, посвященный молекулярным исследованиям и подготовленный А.П. Фоглером (A.P. Vogler, стр. 17-22), несмотря на многочисленные парадоксальные выводы, заслуживает особого интереса. Он представляет собой обобщение всех данных, полученных из сравнения последовательностей нуклеотидов. Автор обсуждает возможности тех или иных генов для решения определенных задач. Например, сравнение последовательностей митохондриальных нуклеотидов [Cytochrome Oxydasa I, Cytochrome b, 16S rRNA] дает хорошие результаты при изучении форм одного рода или близких родов, в то время как ядерная 18S rRNA приемлема для изучения на уровне семейства. Автор располагал информацией о последовательностях нуклеотидов 2 662 видов из 123 семейств (при признании 167 семейств в современной мировой фауне), при этом он отмечает неравномерное распределение имеющихся данных по группам: имеется около 400 видов и подвидов из рода Carabus и более 250 видов из короедов (Scolytidae), в то время как представительство других групп значительно меньше. Для изучения отношений внутри подсемейства Harpalinae с хорошими результатами использовалась 28S rRNA и гены “wingless”. На основании сравнения ДНК делается вывод, например, о монофилетичности как Geadephaga, так и Hydradephaga, причем последние, по мнению автора, трижды осваивали плавание в водной среде (это уже парадокс - см. "Ценная монография по водным жукам Китая"). Во-первых, как можно это утверждать для “монофилетической” группы – не логичнее ли предположить, что разные водные формы монофилетической группы независимо несколько раз вторично утрачивали способность к активному плаванию (особенно если принимать гипотезу водного происхождения подотряда в целом и срастание метакокс с заднегрудью как приспособление именно к плаванию). А.П. Фоглер располагает изученные семейства в оригинальной дендрограмме, в которой подотряд Myxophaga находится ближе к основанию (Archostemata), чем подотряды Adephaga и Polyphaga. Интересны выводы этого автора о “монофилии” Staphyliniformia и Scarabaeiformia, которые согласуются со сравнениями и выводами М. Хансена (Hansen, 1991, 1995). Интересно, что А.П. Фоглер считает, что Scirtoidea следует рассматривать “сестричной” группой по отношению ко всем остальным элатериформным (Elateriformia), а надсемейства Buprestoidea и Dascilloidea более обособленными от 3 других елатериформных надсемейств (Elateroidea, Cantharoidea и Byrrhoidea), причем водные элатериформные семейства по его мнению не “монофилетичны”. Особенно парадоксальными кажутся выводы о том, что надсемейство Cucujoidea является “парафилетичным” по отношению к Cleroidea и многие семейства Cucujoidea попадают между Lymexyloidea и Tenebrionoidea, а также семейства Cryptophagidae, Latrididae, Phalacridae и Corylophidae распределяются среди тенебрионоидов, а семейство Kateretidae и семейство Trogossitidae оказываются в базальном узле надсемейства Cleroidea.
В общем разделе по подотряду Adephaga, подготовленном одним из редакторов и И. Риберой (R.G. Beutel & I. Ribera – стр. 53-55), довольно значительное место отводится сравнению молекулярных данных (по последовательностям ДНК). Здесь же отмечается, что плавунчики (Haliplidae) занимают особое положение как морфологически, так и в силу особенностей строения ДНК, причем по мнению авторов «сестринские» отношения плавунчиков и других адефаг не вполне прослеживаются (“not very well supported”). Они также указывают на особое мнение по положению этого семейства, высказанному А.Г. Пономаренко (1977) и Д.Х. Кавано (Kavanaugh, 1986), к сожалению, не обсуждая его. По-видимому, весьма важным фактом для понимания этой группы является наличие в палеонтологической летописи палеоэндемичного семейства Triaplidae (средний и верхний триас), представленного формами, очень сходными с плавунчиками. Весьма вероятно, что эта вымершая группа филетически связана с современным семейством Haliplidae. Существуют несколько существенных обстоятельств, важных для понимания происхождения и положения этих адефажных семейств. Во-первых, их рецентные виды, как, вероятно, и триасовые, являются, в противоположность другим хищным группам Adephaga, растительноядными. Во-вторых, основание брюшка их имаго отличается от такового у других адефаг неполной консолидацией базальных вентритов. Если принимать более родственные отношения адефаг и миксофаг, а также недавнее описание Meruidae (Spangler & Steiner, 2005), то допущение о том, что плавунчики представляют довольно древнюю и сохраняющую много архаичных черт группу, в том числе и в особенностях трофики (Kirejtshuk, 1991), а именно: альгофагию, что напоминает скорее трофику Meru phyllisae Spangler & Steiner, 2005 и представителей подотряда Myxophaga, чем таковую всех остальных групп Adephaga.
Раздел по плавунцам (Dytiscidae), написанный М. Балке (M. Balke – стр. 90-116), - один из наиболее подробных в книге и наиболее разнообразно иллюстрированный. В нем обращает на себя внимание замечательная схема распределения экологических предпочтений основных родов всех подсемейств (Matinae, Laccophilinae, Coptotomonae, Copelatinae, Hydrodytinae, Hydroporinae, Lancetinae, Colymbetinae, Agabinae и Dytiscinae) и триб, которая сопровождает обзор биономии и экологии семейства, при этом сведения по распространению групп семейства даны лишь в самом общем виде. При описании филогении и таксономии дана таксономическая интерпретация, принимаемая автором, приводится довольно много различных кладограмм и много молекулярной аргументации для филогенетических интерпретаций, но известные исторические данные не привлекаются. Другим по тщательности разбора является раздел по жужелицам (Carabidae) (E. Arndt, R.G. Beutel, K. Will – стр. 119-146), которые достаточно многочисленны и разнообразны и поэтому также разобраны по подсемействам (Paussinae, Gehringiinae, Omophroninae, Cicindelinae, Carabinae, Hiletinae, Loricerinae, Elaphrinae, Megadopinae, Siagoninae, Scaritinae, Trechinae, Harpalinae, Pseudomorphinae и Brachininae). Описание их филогении сводится по существу к сравнению структурных и молекулярных особенностей, выполненному по методологии кладистики без рассмотрения достижений классической карабидологии. Авторы делают некоторые выводы об историческом развитии сем. Carabidae и его филогении, но без каких-либо ссылок на обширные данные по истории семейства. Многие, в том числе почти все небольшие семейства как Adephaga, так и остальных подотрядов, изложены довольно лаконично по определенной редакторами схеме, хотя нередко авторы существенно отклоняются и от этой схемы и от общей таксономической интерпретации, принимаемой редакторами.
В инфраотряд или серию Staphyliniformia редакторы тома не включили надсемейство Scarabaeoidea по той причине, что, по их мнению, эти группы имеют только одну надежную синапоморфию – отсутствие метакоксальной пластинки, а остальные особенности, которые позволяют допускать гомологичность, не могут быть интерпретированы как следствие их общего происхождения. Довольно подробно написан раздел по водолюбам (Hydrophilidae) (M. Archangelsky, R.G. Beutel & A. Kumarek - стр. 158-183), которые прекрасно иллюстрированы и разобраны до уровня подсемейств (Helophorinae, Epimetopinae, Georessinae, Hydrochinae, Spercheinae, Horelophinae, Horelophopsinae, Hydrophilinae, Hydrophilinae и Sphaeridiinae), а последние два подсемейства - до триб, а именно Hydrophilinae: Sperchopsini, Berosini, Chaetarthriini, Anacaenini, Laccobiini и Hydrophilini, а также Sphaeridiinae: Rygmodiini, Coelostomatini, Protosternini, Omicrini, Megasternini и Sphaeridiini. При этом особенности разнообразной биономии взрослых жуков и личинок обсуждаются достаточно детально. Карапузики (Histeridae) (P.W. Kovarik & M.S. Caterino - стр. 190-121) разобраны по подсемействам (Abraeinae, Chlamydopsinae, Hataeriinae, Histerinae, Niponiinae, Onthophilinae, Saprininae, Tribalinae и Trypanaeinae) и трибам, для каждой из которых приводится список родов и указанием распространения каждого из родов. Чрезвычайно подробно описаны разнообразные биономия и экологические приуроченности карапузиков. Семейства Agyrtidae (стр. 261-269) и Leiodidae (стр. 269-280), подготовленные А.Ф. Ньютоном (A.F.Newton), достаточно лаконичны и информативны, с подробными обсуждениями филогенетических схем, выработанных на основании кладистического анализа. Раздел по семейству Scydmaenidae (S.T.O. Keepe - стр. 280-288) включает в том числе полный список родов современной фауны с указанием распространения для каждого, а также атлас взрослых жуков - представителей основных групп семейства. Обзор огромного по численности и многообразию семейства Staphylinidae, объединяющего по подсчетам автора 47 744 вида, занимает довольно значительную часть книги (M.K. Thayer – стр. 296-344). Этот обзор снабжен списком подсемейств (включая подсемейство Pselaphinae в омалиинной линии) с указанием числа родов и числа видов в каждом из них, замечательным и подробным описанием особенностей биономии и экологии различных групп, интересной схемой по типам трофики в различных подсемействах, а также схемой по распределению этих подсемейств в различных экологических условиях и различных географических зонах. Довольно большую и важную часть этого раздела представляет подробное обсуждение филогенетических отношений, которое, однако, не сопровождается ссылками на палеонтологические материалы, а также список литературных источников (14 страниц).
Один из самых больших и интересных разделов посвящен инфраотряду или серии Scarabaeiformia, который или которую обособленно признают авторы раздела (C.H. Scholtz & V.V. Grebennikov - стр. 345-425), несмотря на то, что многими эта самостоятельность оспаривается. В отличие от многих других групп, для скарабеиформных, включающих только одно надсемейство (Scarabaeoidea), достаточно хорошо используются данные по вымершим фаунам. Из различных гипотез авторы предпочитают гипотезу о трофической зависимости предков этой группы от грибов в детрите, обосновывая свое мнение в том числе и приводимой таблицей трофики современных групп, заимствованной из публикации K.Л. Шольца и С.Л. Чоуна (Scholtz & Chown, 1995). Однако представляется, что гипотезы о связях предков скарабеоидных с разлагающейся древесиной (Richter, 1958; Iablokoff-Khnzoryan, 1977 и т.д.) следует признать более оправданными, а образ жизни и способ питания рогачей (Lucanidae) и сахарных жуков (Passalidae) в определенной мере сходным с таковыми предков, хотя, по-видимому, образ жизни последних существенно изменен в связи с очевидно вторично развившейся социальностью. Более тесная связь обеих активных фаз жизненного цикла с разлагающейся древесиной вероятнее других типов отношений, допустимых для предков. Произошло это, весьма вероятно, в течение средней юры (см. "Систематический список ископаемых жуков подотряда Scarabaeina (2-я часть)") и по крайней мере в верхней юре скарабеоидные уже достигли значительного представительства в фауне. Дальнейшие изменения трофики и образа жизни были связаны с тем, что, вероятно, сперва взрослые жуки, а затем и личинки осваивали другие среды обитания и субстраты питания, при этом разлагающаяся органика растительного происхождения оставалась предпочитаемым веществом для питания личинок. В определенной мере в питании личинок кравчиков (Lethrus spp.) и личинок навозников (Aphodiinae и Scarabaeinae), питающихся не до конца переработанной пищей травоядных, можно проследить исходную приуроченность предков группы. Кроме того, в этом разделе детально разобрана аргументация филогенетических связей скарабеоидных, при этом довольно корректно разбираются и точки зрения, не разделяемые авторами. Если общая часть Scarabaeiformia дана с большой детализацией и большим числом иллюстраций, то обзоры семейств – достаточно конспективны, а собственно пластинчатоусые (Scarabaeidae) сопровождаются обзорами всех включенных в него подсемейств.
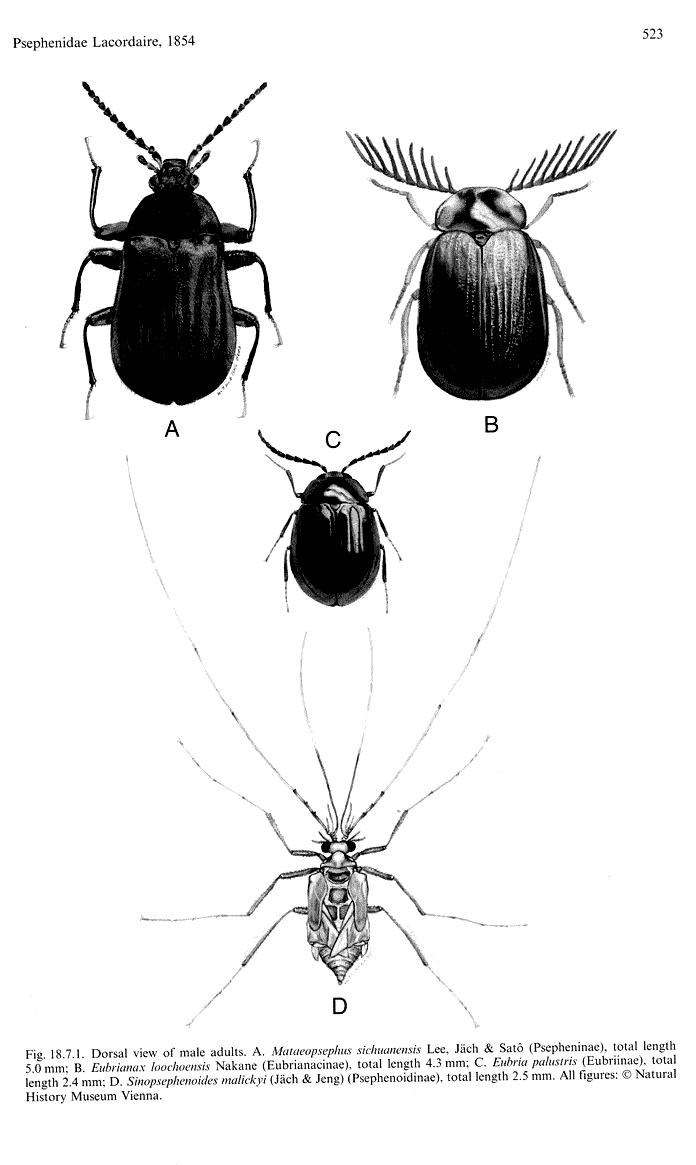
Та часть семейств Elateriformia, которая вошла в первый том, обсуждаются в основном довольно сжато, хотя почти каждое семейство сопровождается отличными иллюстрациями. Вместе с тем разделы по семействам Elmidae (стр. 471-496) и Dryopidae (стр. 496-508), подготовленные Я. Кодадой и М.А. Йехом (J. Kodada & M.A. Jaech), достаточно подробны и хорошо иллюстрированы отличными фотографиями особенностей строения взрослых жуков и личинок, причем раздел по первому семейству состоит по существу из двух разделов, посвященных подсемейству Larinae (Potamophilinae) и подсемейству Elminae. Оба раздела имеют список всех современных родов с указанием числа видов и общего распространения для каждого, подробное описание биономии взрослых жуков и особенно личинок, но сравнительно небольшую часть, в которой обсуждается филогения и таксономия. Это представляется достаточно оправданным из-за недостатка материалов по ископаемым фаунам этих групп. Семейство Psephenidae (C.-F. Lee, M.A. Jaech & R.G. Beutel – стр. 521- 533), имеющее в современной фауне всего 272 вида и разделенное на 4 подсемейства (Eubrianacinae, Eubriinae, Psepheninae и Psephenoidinae), также снабжено подробным списком родов с указанием числа видов и общего распространения для каждого, отличными иллюстрациями, подробным описанием биономии взрослых жуков и особенно личинок, но часть, посвященная филогении, включает детальное кладистическое сравнение отдельных групп, входящих в это семейство, а также сравнение их с другими семействами, в частности, с семействами Elminae и Lutrochinae.
Трудно подготовить столь емкую книгу без изъянов. Бросается в глаза то, что редакторы обычно старались включать ссылки на последние источники, предпочитая при этом компилятивную литературу. В результате чего можно обнаружить многочисленные неточности в интерпретациях или даже двусмысленности в ссылках. Особенно это касается вопросов эволюции, филогении и палеонтологии в общих разделах. Редакторы, очевидно, отдавали предпочтение кладистическим интерпретациям последних лет, порой отмечая, что остальное, что не вписывается в их схемы, и не заслуживает упоминания. У читателя этих разделов возникает впечатление, что почти все достижения колеоптерологии стали возможными лишь благодаря заслугам немногих специалистов, работающих при вдохновляющем влиянии кладистической идеологии. В качестве характерных примеров можно привести следующие. Р.Г. Бейтель (R.G. Beutel) указывает, что принятые в книге филогенетические отношения жесткокрылых и нейроптероидных (Coleoptera, Neuropterida=Neuropteroidea) были предложены В. Хеннигом (Hennig, 1969), Миколайтом (Mickoleit, 1973) и другими (стр. 1). В действительности, гипотеза, которую принимают редакторы, формировалась в первую очередь благодаря публикациям Ламера (Lamеere, 1917), А.В. Мартынова (1925, 1938) и Р.А.Кроусона (Crowson, 1955, 1960). Редакторы со ссылкой на публикацию Р.Г. Бейтеля и Ф. Хааса (Beutel & Haas, 2002) утверждают, что важнейшей особенностью полифаг является наличие цервикальных склеритов (стр. 152), однако при этом не упоминаются размышления, например, Р.А. Кроусона (Crowson, 1975), о связи присутствия этих склеритов у разных нейроптероидных и у полифаг среди жуков, а также о связи их отсутствия у представителей других подотрядов жуков со степенью подвижности головы и характером соединения головы с переднегрудным сегментом.
Довольно странная особенность мнения редакторов есть игнорирование сведений об ископаемых из-за многочисленных пробелов в них и, с другой стороны, создавать "филогению" только на основании данных по современным группам без учета того, что реальная филогения едва ли была линейным процессом и поэтому формы корреляций между современными видами и группами могут сильно отличаться от таковых в прошлом. Да и группы были другими, и жили они в других условиях. Старания редакторов скорее напоминают попытки "восстановить филогению" тетрапод, отбросив все исторические данные о вымерших фаунах этой группы. Исторические факты, которыми оперируют редакторы, порой могут ввести в заблуждение читателя, особенно в связи с констатацией того, что редактор признается в том , что именно он (Beutel, 1997) сформулировал «синапоморфии» и предложил «сестринские отношения» основных групп жесткокрылых (заметим: при этом почти не учитывая палеонтологическую литературу, но упоминая, что палеонтологические данные нельзя надежно таксономически интерпретировать – например на стр. 6: “not sufficient for a reliable systematic placement”). К примеру, апоморфией чекардоколеид рассматривается «непарный заостренный яйцеклад» (“ovipositor unpaired, acuminate”: стр. 2), в то время как палеоколеоптерологи предполагают, что у этого семейства самки, быть может, сохраняли наружный яйцеклад (плезиотипическая черта не только для жуков, но и для нейроптероидных), который в дальнейшем у самок потомков он полностью погрузился внутрь брюшка (Пономаренко, 1969). Со ссылкой на палеонтологов (но не палеоколеоптерологов) Э. Росса и Е. Яржембовского (Ross & Jarzembovski, 1993) в книге утверждается, что, в отличие от предшествующих жуков, представители Taldycupedidae приобретают выраженные эпиплевры (стр. 3). Вместе с тем уже самые древние чекардоколеиды (Tshekardocolidae) характеризуются перегибом по субкостальной жилке (SC), т.е. эпиплеврами, о которых пишут палеоколеоптерологи (в том числе и в книге Пономаренко, 1969) и которые являются важной диагностической особенностью отряда в целом. Редактора обсуждаемого тома считает, что А.Г. Пономаренко в вышеупомянутой книге утверждал, что семейство Catiniidae можно филетически связать как предков (“stemlineage”) с подотрядом Myxophaga (стр. 43), при этом редактор высказывает сомнение в обоснованности такой связи потому, что якобы А.Г. Пономаренко не дает обоснования для этого "a specific justification for this placement". Но указания на такую связь в той монографии, на которую ссылается редактор, вовсе нет. Редактор считает, что А.Г. Пономаренко рассматривал схизофориформных архостемат как водные группы по причине "гладких покровов, обтекаемой формы тела, крючковидных структур на ногах для возможного прикрепления или пребывания на кодных растениях" ("smooth surface, stream-lined body shape, hook-like structures of fore leg for possible attachment to or location on water plant" - стp. 7), и "особенно потому, что многие эти ископаемые найдены в пресноводных отложениях" ("especially by the fact, that many of these fossils were found in freshwater deposits" - стp. 7). За эти вышеуказанные черты отвечает редакто, однако мнение Пономаренко в основном основывалось на присутствии "схизы" ("schiza") у групп Schizophoriformia, которая интерпретировалась как адаптация к пребыванию в воде для фиксации надкрылий и боков брюшка, и для защиты пространства под надкрыльями от проникновения воды. Вероятно, следует указать, что большинство наземных ископаемых доступны благодаря отложению органики в водоемах и изучению их в лакустриновых известняках. Однако было бы неверным считать все попавшее в этот субстрат как относящееся к обитанию в воде. Со ссылкой на палеонтолога (но не палеоколеоптеролога) Ч. Лабандейры (Labandeira, 1994) редактор указывает на появление семейства Cupedidae в нижней перми (“Sakmarian”), однако палеоколеоптерологи и колеоптерологи, занимающиеся эволюцией и филогенией, знают, что это семейство отмечается не ранее триаса, а Р.А. Кроусон (Crowson, 1981) связывает появление этой группы с наиболее ранними следами бурения древесины. Удивительна фраза редактора: «Трансформации признаков личиночных стадий вероятно играли несущественную роль в ранней эволюции жуков» (стр. 6: “Character transformations of immature stages have probably played a minor role in early evolution of Coleoptera”). Список такого рода неточностей можно, к сожалению, продолжить.
Нельзя не отметить, что вышедший учебник является довольно значительным обобщением огромного материала, который получен исследователями предшествующего столетия. Она отражает представление о многих группах с точки зрения специалистов, придерживающихся популярного в последние десятилетия кладистичекого подхода в систематике и филогенетике, при этом нередко игнорируются все остальные точки зрения. В этой связи настоящая книга не может остаться не замеченной и должна рассматриваться как этапная в развитии представлений об этом отряде насекомых. Редакция сайта желает редакторам издания успехов в подготовке следующего тома, при этом бы хотелось бы видеть в следующем томе многообразие подходов и различные точки зрения, а также корректное использование данных палеонтологической летописи.
Большинство упоминаемых мной страниц воспроизведены в расположенной выше аннотации А.Л.Лобанова на рецензируемый том. Ниже я привожу только несколько отсутствующих там или хуже отсканированных страниц.
Январь 2006 г.
Последнее обновление: июль 2006 г.

|

|
|