Морфологические исследования
Уточнения к определительной таблице видов рода Aedes Meigen фауны Северо-Запада России по гениталиям самцов (Diptera, Culicidae)
А.В. Халин
По материалам статьи: А.В. Халин. Уточнения к определительной таблице видов рода Aedes Meigen фауны Северо-Запада России по гениталиям самцов (Diptera, Culicidae) // Энтомологическое обозрение 2008, том ##, вып. #. С. ###-###.
Проведенные нами исследования (Халин, 2005, 2006, 2007, 2008) показали, что признаки трехмерной формы структур тела комаров актуальны для определения видов рода Aedes Meigen*. Обширный материал для диагностики представляют признаки генитального аппарата самцов (рис. 1), однако точные данные об их сложной объемной структуре можно получить лишь при исследованиях под разными ракурсами. Нами было показано, что строение генитального аппарата самцов кровососущих комаров (сем. Culicidae) значительно сложнее, чем представляли исследователи (Edwards, 1932; Штакельберг, 1937; Carpenter, LaCasse, 1953; Belkin, 1962; Mohrig, 1969; Гуцевич и др., 1970; Knight, Laffoon, 1971; Becker et al., 2003). Изучение методами световой микроскопии (СвМ) многих структур гениталий самцов затруднено, их объемная конфигурация оказывается более доступной для исследования под растровым электронным микроскопом (РЭМ).
*В настоящей публикации автор использует классификацию Эдвардса (Edwards, 1932), согласно которой таксоны Finlaya, Rusticoidus и Ochlerotatus рассматриваются в качестве подродов рода Aedes. Представления об объеме рода Aedes других исследователей [например, Рейнерта (Reinert, 2000; Reinert et al., 2004, 2006, 2008)] были изложены ранее (Халин, 2005, 2007; Халин, Горностаева, 2008)

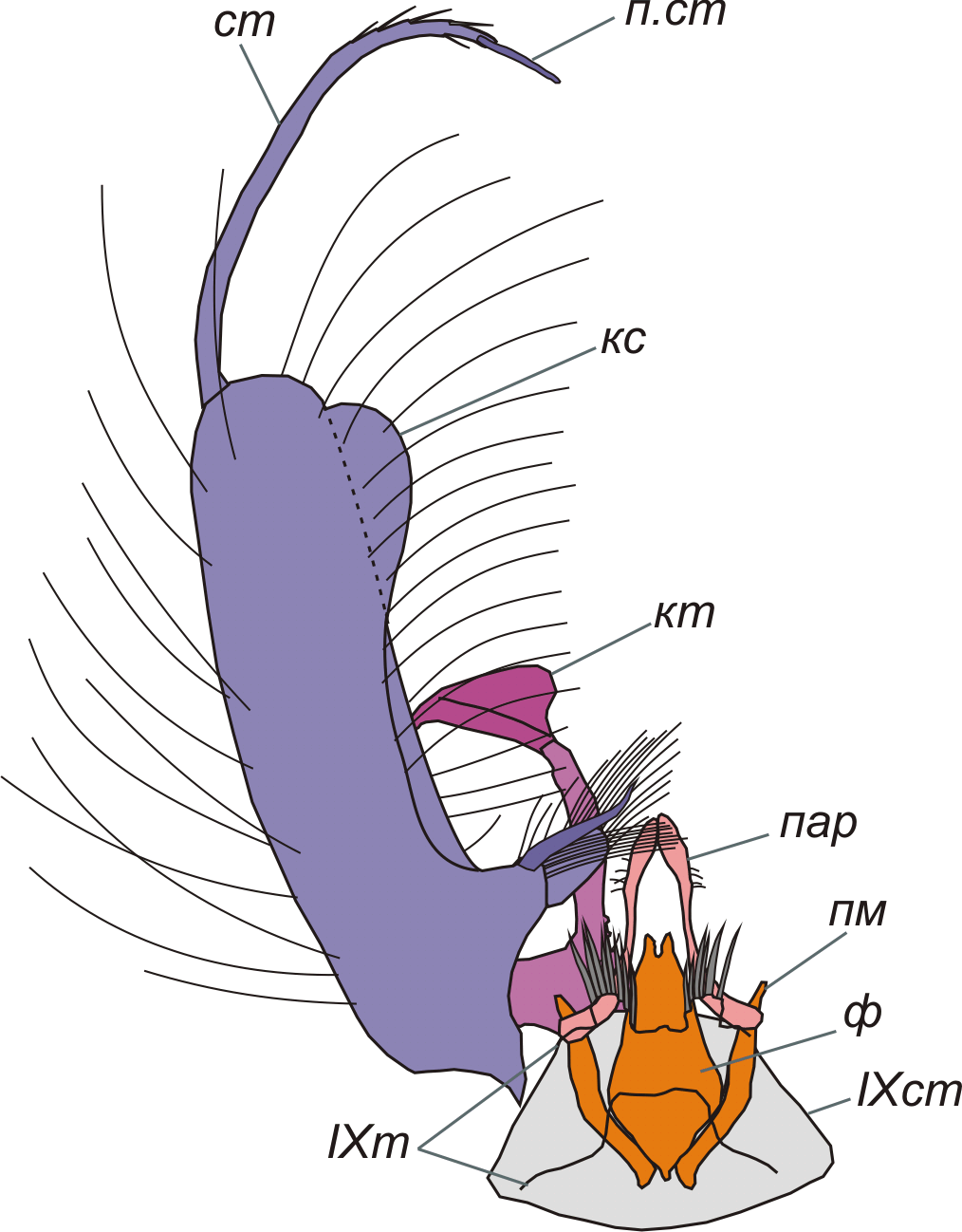
Рис. 1-A. Aedes Meigen, гениталии самца снизу, схема (IX тергит изображен прозрачным). кс – коксит; кт – класпет; пар – парапрокт; пм - парамера; п.ст - придаток стиля; ст – стиль; ф - фаллосома; IX ст и IX т – 9-й стернит и тергит брюшка.
|

Рис. 1-Б. Aedes cantans (Meigen). Гениталии самца снизу под СвМ. Обозначения, как на рис. 1-А.
|
Основу настоящей работы составляет детальное изучение методами РЭМ и СвМ генитального аппарата самцов рода Aedes. Результаты исследования позволили скорректировать определительные таблицы видов данного рода из монографий Беккера с соавторами (Becker et al., 2003), Морига (Mohrig, 1969) и работы Гуцевича и Дубицкого (1981), а также составить ключ для видов рода Aedes фауны Северо-Запада России (Калининградская, Псковская, Ленинградская, Новгородская, Мурманская, Архангельская и Вологодская области, а также Республики Карелия и Коми). Новые и уточненные данные по диагностике видов, отмеченные нами, выделены в тексте таблицы полужирным шрифтом. В определительную таблицу включено 29 видов из 5 подродов рода Aedes, находки которых возможны на территории Северо-Запада России согласно литературным данным (Халин, Горностаева, 2008). Некоторые виды [например, Aedes (Finlaya) geniculatus (Oliver), A. (Rusticoidus) rusticus (Rossi)] включены в таблицу условно, их распространение на территории Северо-Запада России требует подтверждения.
За ценные консультации автор выражает благодарность С. Г. Медведеву (Зоологический институт РАН). Также я глубоко признателен ведущему инженеру Т. К. Цогоеву за помощь при работе с РЭМ.
Работа выполнена на базе коллекции Зоологического института РАН (ЗИН РАН) (УФК ЗИН рег. № 2-2.20) и на основании контракта с Роснаукой «02.452.11.7031 (2006-РИ-26.0/001/070), при поддержке Российского фонда фундаментальных исследований (грант 08-04-00216-а) и гранта Президента РФ поддержки научных школ № НШ-1664.2003.4.
Материал и методика
Работа выполнена в Лаборатории паразитологии ЗИН РАН. Для изучения генитального аппарата были использованы комары, собранные автором в Ленинградской области и на юге Украины в течение 2002-2005 гг. (спиртовой материал), а также материалы фондовой коллекции ЗИН РАН.
Генитальный аппарат самцов изучен нами у 29 видов рода Aedes из подродов Aedes, Aedimorphus Theobald, Finlaya Theobald, Rusticoidus Shevchenko et Prudkina и Ochlerotatus Lynch Arribalzaga. 50 экз., относящихся к 21 виду, исследованы под РЭМ; более 300 экз., принадлежащих к 29 видам рода Aedes, изучены при помощи СвМ.
Исследованы следующие виды:
Aedes (Aedes) cinereus Meigen, 1818;
A. (A.) rossicus Dolbeskin, Gorickaja et Mitrofanova, 1930;
A. (Aedimorphus) vexans vexans (Meigen, 1830);
A. (Finlaya) geniculatus (Olivier, 1791);
A. (Ochlerotatus) annulipes (Meigen, 1830);
A. (O.) behningi Martini, 1926;
A. (O.) cantans (Meigen, 1818);
A. (O.) caspius (Pallas, 1771);
A. (O.) cataphylla Dyar, 1916;
A. (O.) communis (De Geer, 1776);
A. (O.) cyprius Ludlow, 1920;
A. (O.) diantaeus Howard, Dyar et Knab, 1913;
A. (O.) dorsalis (Meigen, 1830);
A. (O.) euedes Howard, Dyar et Knab, 1913;
A. (O.) excrucians (Walker, 1856);
A. (O.) flavescens (Muller, 1764);
A. (O.) hexodontus Dyar, 1916;
A. (O.) impiger (Walker, 1848);
A. (O.) intrudens Dyar, 1919;
A. (O.) leucomelas (Meigen, 1804);
A. (O.) mercurator Dyar, 1920;
A. (O.) nigrinus (Eckstein, 1918);
A. (O.) nigripes (Zetterstedt, 1838);
A. (O.) pionips Dyar, 1919;
A. (O.) pullatus (Coquillett, 1904);
A. (O.) punctor (Kirby, 1837);
A. (O.) riparius Dyar et Knab, 1907;
A. (O.) sticticus (Meigen, 1838);
A. (Rusticoidus) rusticus (Rossi, 1790).
Для исследований под РЭМ главным образом использовались особи, хранящиеся в 70% водном растворе спирта. Последние сегменты брюшка комара отделялись и обрабатывались водным раствором щелочи, затем промывались дистиллированной водой, обезвоживались в спиртах возрастающей концентрации и в ацетоне. Обработка щелочью производилась с целью очистки поверхности структур от загрязнений; заметной деформации покровов при проведении данной процедуры не наблюдалось. Просушка объекта осуществлялась в углекислоте методом «критической точки». После этого объект наклевался на специальный столик при помощи двухстороннего скотча и напылялся платиной.
Помимо методов РЭМ нами использовалась СвМ, совместимая с компьютером. Изображение со светового микроскопа («Leica DM LS» и «LeicaDM5000B») передавалось на персональный компьютер «Pentium IV» и «Core™2» посредством фотокамер «Cannon Coolpix 4500» или «Leica DFC320».
Определительная таблица по самцам
1. Стиль простой, удлиненный конусовидный или эллипсоидной формы, не разделен на 2 ветви. Место сочленения стиля с кокситом расположено на вершине коксита (рис. 1). Щупики самцов приблизительно равны по длине хоботку………2
Стиль сложной формы, состоит из двух ветвей: короткой, занимающей дорсо-медиальное положение, и более длинной, расположенной вентро-латерально. Место сочленения стиля с кокситом находится на дорсальной поверхности коксита. Расстояние от вершины коксита до места сочленения стиля составляет от 1/4 до 1/3 длины коксита. Щупики самцов в несколько раз короче длины хоботка
………подрод Aedes, 25
2 (1). Класпеты имеют вид парных удлиненных выростов, сочлененных с 9-м стернитом, разделены на стволик и крыло (рис. 2, 3, 4). Стиль равномерно сужается к вершине. Придаток стиля расположен на вершине стиля (рис. 5)………3

Рис. 2. Aedes diantaeus Howard, Dyar et Knab, коксит снизу, бальзамный препарат (фондовая коллекция ЗИН РАН, Ленинградская обл., Громово, 22.VI.1971).
б. б. и в. б - базальная и вершинная бородавки коксита, п. щ - пучок щетинок коксита.
|

Рис. 3. Aedes diantaeus Howard, Dyar et Knab, бородавка коксита и класпет снизу, бальзамный препарат (фондовая коллекция ЗИН РАН, Ленинградская обл., Громово, 22.VI.1971). д. щ и пр. щ - дистальные и проксимальная щетинки базальной бородавки коксита, к. кл и с. кл - крыло и стволик класпета.
|

Рис. 4. Aedes diantaeus Howard, Dyar et Knab, класпет сверху под РЭМ.
|

Рис. 5. Aedes diantaeus Howard, Dyar et Knab, дистальная часть вальвы снизу, бальзамный препарат (фондовая коллекция ЗИН РАН, Санкт-Петербург, лесопарк Александринино, 15.VI.1970). п. ст – придаток стиля.
|
Класпеты не разделены на крыло и стволик, имеют вид колбовидных образований. На вершине каждого класпета расположено около 20 щетинок различной длины (некоторые из них превышают длину класпета). В дистальной части данные щетинки крючковидно изогнуты. Стиль эллипсоидной формы, немного расширяется к вершине. Придаток стиля причленяется на дополнительном выступе, расположенном на небольшом расстоянии от вершины стиля (приблизительно 1/5 длины стиля)………A. (Aedimorphus) vexans
3 (2). Коксит конусовидной формы без выраженной вершинной бородавки. Базальная бородавка коксита имеет вид едва намеченной лопасти у основания коксита (рис. 6). Крыло класпета серповидной формы, примерно равно по длине стволику класпета………A. (Finlaya) geniculatus
.JPG)
Рис. 6. Aedes geniculatus (Olivier), базальная бородавка коксита снизу под СвМ.
|
Коксит обычно сложной формы с более или менее развитой базальной бородавкой в виде выступа (рис. 2). Также как правило, имеется и вершинная бородавка коксита (более или менее выступающая лопасть). Крыло класпета клинковидной, треугольной, копьевидной или иной формы (рис. 7-17). Его длина короче стволика класпета, реже длинее его………4

Рис. 7. Aedes diantaeus Howard, Dyar et Knab, крыло класпета сзади под РЭМ. о. крл – «окошечко» крыла класпета.
|

Рис. 8. Aedes communis (De Geer), крыло класпета сбоку под РЭМ.
|

Рис. 9. Aedes pullatus (Coquillett), крыло класпета сзади под РЭМ.
|

Рис. 10. Aedes communis (De Geer), крыло класпета сбоку под СвМ.
|

Рис. 11. Aedes intrudens Dyar, крыло класпета сзади под РЭМ.
|

Рис. 12. Aedes mercurator Dyar, крыло класпета сзади под РЭМ. р. крл – «прозрачное расширение» крыла класпета; рук. крл – «рукоятка» крыла класпета.
|

Рис. 13. Aedes flavescens (Muller), крыло класпета сбоку под РЭМ. ст. крл – «стержень» крыла класпета.
|

Рис. 14. Aedes punctor (Kirby), крыло класпета сбоку под РЭМ.
|

Рис. 15. Aedes sticticus (Meigen), крыло класпета сбоку под РЭМ.
|

Рис. 16. Aedes excrucians (Walker), крыло класпета сбоку под РЭМ.
|

Рис. 17. Aedes annulipes (Meigen), крыло класпета сбоку под СвМ.
|
4 (3). Базальная бородавка коксита имеет вид раздвоенного выступа (рис. 18). На задней поверхности данных выступов расположены изогнутые и уплощенные кутикулярные выросты, в несколько раз превышающие длину бородавки. Некоторые из них покрыты микротрихиями. Придаток стиля S-образно изогнут. На крыле класпета имеются гребни, расположенные почти перпендикулярно продольной оси пластинки крыла. Фаллосома состоит из медиальной удлиненной склеротизованной части и 2 латеральных долей, несущих на вершине зубцы, в базальной части – латеральные крыловидные выступы……… A. (Rusticoidus) rusticus

Рис. 18. Aedes rusticus (Rossi), базальная бородавка коксита снизу под СвМ. упл. пр - уплощенные придатки базальной бородавки коксита.
|
Базальная бородавка коксита различного строения, но из кутикулярных образований на ней могут присутствовать только щетинки (в том числе шиповидные или немного уплощенные) и микротрихии (рис. 19-32). Придаток стиля прямой или серповидно изогнутый. На крыле класпета, если имеются гребни или борозды, то они ориентированы большей частью продольно оси крыла (рис. 8, 10, 17). Фаллосома цельная, более или менее овальной формы (на заднем крае с выемкой различной глубины, рис. 33)……… подрод Ochlerotatus, 5

Рис. 19. Aedes cantans (Meigen), базальная бородавка коксита снизу под РЭМ. кр. щ, н. щ, у. щ и ш. щ - крупные, небольшие, утолщенные и шиповидные щетинки базальной бородавки коксита.
|

Рис. 20. Aedes nigripes (Zetterstedt), основание базальной бородавки коксита снизу под СвМ.
|

Рис. 21. Aedes mercurator Dyar, базальная бородавка коксита снизу под РЭМ. дор. п - дорсальная поверхность коксита, п. кр - передний край базальной бородавки коксита.
|

Рис. 22. Aedes diantaeus Howard, Dyar et Knab, базальная бородавка коксита изнутри под РЭМ. д. кр и в. кр – дорсальный и вентральный края базальной бородавки коксита.
|

Рис. 23. Aedes flavescens (Muller), базальная бородавка коксита снизу под РЭМ. вн. п - внутренняя поверхность коксита.
|

Рис. 24. Aedes behningi Martini, базальная бородавка коксита снизу под РЭМ.
|

Рис. 25. Aedes caspius (Pallas). Базальная бородавка коксита сзади под РЭМ; з. п – задняя поверхность бородавки; микр – микротрихии.
|

Рис. 26. Aedes caspius (Pallas). основание шиповидных щетинок базальной бородавки сзади под РЭМ.
|

Рис. 27. Aedes caspius (Pallas). Базальная бородавка коксита снизу под РЭМ. п. п – передняя поверхность бородавки; ш. щ – шиповидная щетинка базальной бородавки.
|

Рис. 28. Aedes caspius (Pallas). Шиповидные щетинки базальной бородавки спереди под РЭМ.
|

Рис. 29. Aedes communis (DeGeer), базальная бородавка коксита снизу под РЭМ. к. щ - крючковидно изогнутые щетинки базальной бородавки коксита.
|

Рис. 30. Aedes pullatus (Coqillett). Базальная бородавка коксита изнутри под РЭМ. уп. щ - уплощенные щетинки базальной бородавки коксита.
|

Рис. 31. Aedes sticticus (Meigen), базальная бородавка коксита снизу под РЭМ. пер - перетяжка базальной бородавки коксита.
|

Рис. 32. Aedes excrucians (Walker). Базальная бородавка коксита изнутри под РЭМ.
|

Рис. 33. Aedes diantaeus Howard, Dyar et Knab, эдеагус снизу. Бальзамный препарат (фондовая коллекция ЗИН РАН, Хабаровский край, Большехехцирский заповедник, 20.VII.1976). б. п – базальная пластинка.
|
5 (4). Базальная бородавка коксита c 3 утолщенными щетинками, 1 из которых находится в проксимальной части бородавки и направлена внутрь, 2 – в дистальной, направлены назад и внутрь (рис. 3, 22). Плоскость крыла класпета ориентирована перпендикулярно стволику (рис. 4). Фаллосома сильно склеротизована с заметными элементами разделения на латеральные доли (рис. 33)………группа видов “intrudens”, 6
Базальная бородавка коксита c 1 утолщенной щетинкой шиповидного типа или без нее (рис. 19-21, 23, 24, 29-32). Также на базальной бородавке может присутствовать 2-я, дополнительная шиповидная щетинка, заметно уступающая по размеру 1-й (рис. 25-27). Плоскость крыла класпета может быть расположена под различным углом к стволику. Фаллосома склеротизована в различной степени, на заднем крае может присутствовать небольшая выемка, однако явных признаков разделения на латеральные доли нет………8
6 (5). Все 3 утолщенные щетинки базальной бородавки коксита шиповидного типа (рис. 3, 22). Стволик класпета около середины на внутренней поверхности с хорошо различимым пальцевидным выростом, несущим щетинку (рис. 4). На крыле класпета имеются слабо склеротизованные участки в форме овальных окошек (рис. 3, 7)………7
Базальная бородавка коксита с 1 шиповидной щетинкой в проксимальной части и 2 уплощенными щетинками в дистальной части (рис. 30). Стволик класпета немного проксимальнее середины расширен и изогнут; выступ на внутренней поверхности, если имеется, едва различим. Крыло класпета удлиненное, с сильно загнутой вершиной, направленной дорсально и вперед; пластина крыла неровная, на дорсальной поверхности имеется заметное продольное углубление. На крыле класпета слабо склеротизованных участков в форме овальных окошек нет (рис. 9)………A. pullatus
7 (6). На вентральной поверхности коксита в его дистальной половине расположен пучок длинных изогнутых щетинок, направленных внутрь (рис. 34). Стволик класпета с выростом на внутренней поверхности, расположенным дистальнее середины (рис. 4). Крыло класпета в форме равностороннего треугольника с загнутыми углами, ориентировано перпендикулярно стволику класпета, вершина направлена внутрь. Слабо склеротизованные участки в форме овальных окошек на крыле класпета хорошо различимы (рис. 3, 7)………A. diantaeus

Рис. 34. Aedes diantaeus Howard, Dyar et Knab, внутренняя поверхность коксита под РЭМ. вен. п - вентральная поверхность коксита, м - мембранозный участок внутренней поверхности коксита, о. к - основание коксита.
|
Пучка длинных изогнутых щетинок на вентральной поверхности коксита нет. Лишь небольшая субапикальная часть коксита с густыми щетинками, направленными большей частью дистально. Вырост стволика класпета находится на его внутренней поверхности проксимальнее середины. Крыло класпета удлиненное, формы птичьей головы с изогнутой вершиной; ориентировано перпендикулярно стволику, направлено вершиной дорсально и немного вперед. На крыле класпета слабо склеротизованные участки типа окошек едва заметны (рис. 11)………A. intrudens
8 (5). На базальной бородавке коксита помимо 1 шиповидной щетинки имеется 2-я дополнительная шиповидная щетинка, заметно уступающая по размеру 1-й, расположенная ближе к дорсальному краю бородавки (рис. 25-27). Базальная бородавка в виде трутового гриба, ориентирована почти перпендикулярно продольной оси коксита (рис. 27). Лапки со светлыми кольцами, опоясывающими дистальную и проксимальную части каждого членика………9
Базальная бородавка коксита лишь с 1 шиповидной щетинкой (рис. 19, 21, 23, 29, 31) или без нее (рис. 20, 24, 32). Дополнительной шиповидной щетинки, расположенной ближе к дорсальному краю бородавки, нет; единственная шиповидная щетинка сама занимает крайнее дорсальное положение на базальной бородавке. Базальная бородавка различной формы. Лапки без светлых колец или светлые кольца опоясывают только проксимальные части члеников………10
9 (8). Главная и дополнительная шиповидные щетинки расположены сравнительно близко друг к другу (расстояние превышает примерно в 2.5 раза диаметр главной шиповидной щетинки, рис. 26)………A. caspius
Шиповидные щетинки удалены друг от друга (расстояние превышает примерно в 3.5-4 раза диаметр главной шиповидной щетинки)………A. dorsalis
10 (8). Вершинная бородавка коксита слабо выражена, имеет вид небольшой лопасти на внутренней поверхности коксита. Базальная бородавка в виде трутового гриба; по краю с рядом щетинок, наиболее крупные из них занимают дорсальное положение. Стволик класпета сильно изогнут в срединной трети, крыло удлиненное, серповидной формы с 1 хорошо различимым латеральным слабо склеротизованным участком. Фаллосома на вершине с 2 изогнутыми зубцевидными выступами, направленными вершинами внутрь………11
Вершинная бородавка коксита хорошо развита, в виде более или менее выступающей лопасти (заметно выступает в промежуток между вальвами на бальзамных микрорепаратах). Базальная бородавка, класпеты и вершина фаллосомы различного строения………12
11 (10). Щетинки коксита, направленные внутрь (расположенные главным образом на вентральной поверхности коксита) сравнительно длинные, некоторые из них почти достигают длины коксита. 3-4 щетинки базальной бородавки коксита, занимающие крайнее дорсальное положение, практически одинакового строения (т.е. шиповидной щетинки, четко выделяющейся на фоне других щетинок, нет, рис. 20). Фаллосома сильно склеротизована. Лопасти 9-го тергита с 14-20 щетинками каждая, причем значительная часть из них (около 10) расположена на дорсальной поверхности лопасти………A. nigripes
Щетинки коксита, направленные внутрь (расположенные главным образом на вентральной поверхности коксита) сравнительно короткие (как правило, около половины длины коксита). На базальной бородавке коксита крайнее дорсальное положение занимает шиповидная щетинка, заметно превышающая по толщине (примерно в 2 раза) расположенные рядом щетинки. Фаллосома слабо склеротизована. Лопасти 9-го тергита с 4-12 щетинками каждая, только 1 или 2 из которых расположены на дорсальной поверхности лопасти, остальные – ее на заднем крае………A. impiger
12 (10). Базальная бородавка коксита с 1 шиповидной щетинкой, выделяющейся на фоне остальных щетинок и занимающей крайнее дорсальное положение среди остальных щетинок бородавки (рис. 19, 21, 23, 29, 31)………13
Базальная бородавка коксита без шиповидных щетинок, четко выделяющихся толщиной по сравнению с остальными щетинками (рис. 24, 32). Таким образом, все щетинки базальной бородавки оказываются примерно одинаковой длины и толщины
………23
13 (12). Крыло класпета на наружной поверхности с несколькими (обычно от 3 до 5) продольными бороздами. Из них, по крайней мере, 1 хорошо заметна, она сильно выступает латеро-каудально, нередко почти достигая заднего края крыла класпета. На внутренней поверхности крыла класпета имеются соответствующие углубления. Таким образом, пластина крыла оказывается гофрированной, что заметно при изучении ее как методами РЭМ, так и СвМ (рис. 8, 10). Крыло класпета расположено в сагиттальной плоскости; имеет удлиненно-треугольную форму, вершина загнута вниз. Стволик класпета длинный (примерно в 2 раза короче коксита), утолщенный в базальной трети (длина более чем в 7 раз превышает толщину у основания), слабо изогнутый, направлен крылом вперед. Задняя поверхность базальной бородавки коксита сильно вогнутая, таким образом, бородавка по форме напоминает корзинку (рис. 29)………A. communis, A. pionips
Крыло класпета, если и с продольными ребрами, то с едва заметными; по крайней мере, снизу оно не выглядит гофрированным. В остальном класпеты могут иметь различное строение. Базальная бородавка коксита различного строения, но ее задняя поверхность если и вогнутая, то не сильно. Таким образом, бородавка не имеет форму корзинки………14
14 (13). Крыло класпета равномерно сильно склеротизовано, слабо склеротизованные участки на нем отсутствуют (рис. 14). Оно имеет фору птичьей головы, его вершина загнута вниз. Пластина крыла практически ровная, не изогнутая; расположена в сагиттальной плоскости, направлена дорсально. Базальная бородавка коксита крупная, расстояние от ее проксимального края до дистального примерно равно длине стволику класпета. Стволик класпета короткий, примерно в 3 раза короче коксита. У основания стволик класпета утолщен, его длина приблизительно в 4 раза превышает толщину у основания. В базальной трети стволик слабо изогнут
………A. punctor, A. hexodontus
Крыло класпета дифференцировано на сильно склеротизованную стволовую часть и 1-2 слабо склеротизованных боковых участков (рис. 12, 13, 15)………15
15 (14). Направленные внутрь щетинки, расположенные дистальнее базальной бородавки на вентральной поверхности коксита, длинные (некоторые достигают длины коксита), обычно перекрываются концами на бальзамных микропрепаратах (рис. 35). Стволик класпета длинный (около половины от длины коксита) сильно изогнут за серединой (дистальный и проксимальный участки образуют угол примерно 70-80°). Вершинная бородавка сильно выступает внутрь (длина немного превышает ширину у середины)………16

Рис. 35. Aedes leucomelas (Meigen), гениталии самца снизу под СвМ. дл. щ - длинные щетинки вентральной поверхности коксита.
|

Рис. 36. Aedes cataphylla Dyar, лопасти IX тергита снизу под СвМ.
|

Рис. 37. Aedes leucomelas (Meigen), лопасти IX тергита снизу под СвМ.
|

Рис. 38. Aedes cataphylla Dyar, лопасти IX тергита снизу под СвМ.
|
Хотя бы некоторые щетинки, расположенные дистальнее базальной бородавки коксита, короткие (как правило, примерно около 1/2 от длины коксита), не перекрываются концами. Стволик класпета различного строения. Вершинная бородавка выступает внутрь не очень сильно (длина менее ширины у середины)………17
16 (15). Щетинки на лопастях 9-го тергита прямые, направлены назад. Лопасти 9-го тергита удалены друг от друга, расстояние между ними примерно равно ширине лопасти (рис. 36, 38)………A. cataphylla
Щетинки на лопастях 9-го тергита изогнутые, направлены назад и наружу. Лопасти 9-го тергита расположены близко друг к другу, расстояние между ними составляет примерно половину от ширины лопасти (рис. 37)………A. leucomelas
17 (15). Базальная бородавка в виде четко обособленного треугольного выступа с более или менее округлыми вершинами, у основания имеется хорошо различимая короткая перетяжка (рис. 31). Внутренняя поверхность бородавки несет небольшие щетинки, их длина составляет менее половины расстояния от дистального до проксимального края бородавки. Более длинные щетинки, а также 1 шиповидная щетинка, находятся у основания бородавки на дорсальной стороне. Вершинная бородавка с широким основанием, доходящим в проксимальном направлении до середины коксита. Класпеты составляют менее половины от длины коксита. Стволик класпета почти прямой, лишь немного изогнут в дорсальном направлении. Крыло класпета составляет немногим более половины длины стволика, удлиненное, направлено дорсально и назад, вершина немного отогнута вперед. На крыле имеются 2 латеральных утонченных участка: хорошо различимый дистальный и едва заметный проксимальный (рис. 15)………18
Базальная бородавка коксита различного строения, однако, четко выраженной перетяжки у основания нет. Вершинная бородавка ограничена дистальной третью коксита. Крыло класпета с 1 латеральным утонченным участком, дистальным
(рис. 12, 13)………19
18 (17). Дистальный угол базальной бородавки слабо закруглен, немного изогнут внутрь. Таким образом, при рассмотрении снизу базальная бородавка имеет форму полумесяца (рис. 31)………A. sticticus
Дистальный угол базальной бородавки более округлый, не изогнут внутрь………A. nigrinus
19 (17). Базальная бородавка коксита имеет сложную форму (рис. 19). Ее дорсальная часть в виде конического выступа, на вершине которого располагается утолщенная шиповидная щетинка с хорошо заметными продольными бороздами. Рядом с ней находится группа удлиненных щетинок, в 3 раза и более уступающих по толщине шиповидной. От вентральной поверхности конического выступа отходит удлиненная лопасть, направленная назад и вентрально. Ее длина превышает ширину при основании, на вершине лопасти расположены утолщенные щетинки. Стволик класпета примерно в 2.5 раза короче коксита, слабо изогнут дорсально. Крыло класпета направлено дорсально………20
Базальная бородавка различного строения, но удлиненная лопасть, направленная назад и вентрально, отсутствует. Стволик класпета различного строения………21
20 (19). Базальная бородавка с сильно удлиненной лопастью, отходящей в вентральном направлении и дугообразно изогнутой, так что вершина направлена назад (рис. 19). Длина лопасти примерно в 2 раза превышает ширину при основании. Крыло класпета треугольной формы, с очень широким латеральным слабо склеротизованным участком. Ширина крыла составляет, по крайней мере, половину длины крыла класпета. Дистальная часть слабо склеротизованного участка нередко волнообразно изогнута и загнута внутрь………A. cantans
Базальная бородавка с менее удлиненной лопастью, отходящей в вентральном направлении и немного назад. Длина лопасти менее чем в 1.5 раза превышает ширину при основании. Крыло класпета удлиненно-треугольной формы, с умеренно широким латеральным слабо склеротизованным участком. Ширина крыла составляет не более 1/4 от его длины………A. riparius
21 (19). Базальная бородавка в виде треугольной лопасти с округлыми вершинами, отходящей от дорсальной поверхности коксита, направленной вентрально. Таким образом, базальная бородавка располагается вдоль продольной оси коксита (рис. 23). Длина бородавки составляет примерно треть длины коксита. В проксимальной части бородавка немного возвышена, дистально ее высота уменьшается. Расстояние от ее проксимального до дистального края превышает расстояние от дорсального края до вентрального. У основания бородавки ближе к ее проксимальному краю на дорсальной поверхности коксита расположена 1 утолщенная шиповидная щетинка (ее длина чуть меньше длины бородавки). На дорсальной поверхности бородавки расположены более короткие (примерно 0.5 от длины бородавки) и более тонкие (0.1 от толщины шиповидной щетинки) слабо изогнутые щетинки. Стволик класпета короткий (менее трети от длины коксита), утолщенный (толщина составляет более ? от длины), почти прямой. Крыло класпета клинковидной формы, изогнуто у основания, с небольшой рукояткой (менее 1/5 от длины крыла), направлено назад и дорсально (рис. 13)………A. flavescens
Базальная бородавка более сложной формы. Ее дорсальная часть ориентирована практически перпендикулярно продольной оси коксита. Вентральная часть бородавки заметно изогнута назад, образуя с продольной осью коксита небольшой угол (рис. 21)………22
22 (21). Латеральное слабо склеротизованное расширение крыла класпета начинается дистальнее середины крыла класпета, в базальной части крыла имеется так называемая «рукоятка крыла класпета». Крыло класпета удлиненное (превышает длину стволика), копьевидное (рис. 12). Крылья класпетов направлены дорсально, перекрещиваются друг с другом, образуют с продольной осью тела угол близкий к 90°. Длина базальной бородавки примерно равна ее ширине, в 3-4 раза менее длины коксита. На базальной бородавке имеются щетинки, составляющие около 1/2 ее длины или немногим более ее. В проксимальной части бородавки на границе дорсальной и внутренней поверхностей коксита имеется небольшой выступ, на котором расположена крупная шиповидная щетинка. Ее длина в 1.5-2 раза превышает длину бородавки, толщина – более чем в 4 раза превышает таковую остальных щетинок бородавки (рис. 21)
………A. mercurator
23 (12). Базальная бородавка в виде хорошо обособленной лопасти на внутренней поверхности коксита, отчетливо выступающей внутрь (рис. 24). Бородавка имеет форму треугольника с округлыми вершинами, ее длина примерно равна ширине и составляет не более 1/4 от длины коксита. Проксимальная и дорсальная части бородавки немного выступают в дорсальном направлении. На дорсальной поверхности бородавки расположены щетинки различного размера, не превышающие длины бородавки. Стволик класпета удлиненный, его длина примерно в 5 раз превышает ширину. У вершины он слабо изогнут, в 2-3 раза короче коксита. Крыло класпета удлиненно треугольной формы, чуть короче стволика, направлено дорсально. На крыле имеется небольшая рукоятка, составляющая примерно 1/4-1/3 от длины крыла………A. behningi
Базальная бородавка в виде слабо обособленной лопасти на внутренней поверхности коксита, почти не выступающей внутрь и направленной большей частью вентрально (рис. 32). Бородавка имеет форму треугольника с округлыми вершинами, ее длина в 2-3 раза превосходит ширину и составляет примерно 1/2 от длины коксита. В дорсальном направлении бородавка не выступает. Поверхность бородавки практически равномерно покрыта небольшими щетинками, длина которых составляет примерно 1/3 от длины бородавки, направленными большей частью вентрально. В проксимальной части бородавки также имеются микротрихии. Стволик класпета сильно изогнутый у середины, его дистальный и проксимальный участки составляют угол около 90°. Длина стволика класпета составляет примерно 1/2 от длины коксита. Крыло класпета в форме клинка, направлено дорсально, короче стволика примерно в 1.5 раза (рис. 16, 17). Четко выраженной рукоятки крыла класпета нет………24
24 (23). Стволик класпета составляет примерно половину длины коксита, к вершине немного утолщается. Крыло класпета (рис. 17) примерно в 2 раза короче стволика, с 2 или 3 хорошо заметными волнообразно изогнутыми продольными ребрами, сходными с таковыми A. communis, но слабее выступающими. В дистальной трети крыло класпета расширено сильнее, чем в проксимальной………A. annulipes
Стволик класпета к вершине заметно сужается, толщина стволика у основания примерно в 2 раза превышает таковую у вершины. Крыло класпета (рис. 16) примерно в 1.5 раза короче стволика, сужается равномерно к вершине, проксимальный край латерального расширения, как правило, с зубцом………A. excrucians, A. euedes
25 (1). Вентро-латеральная ветвь стиля на вершине двураздельная………A. cinereus
Вентро-латеральная ветвь стиля на вершине зазубрена, не раздвоена………A. rossicus
Замечания по диагностике отдельных видов рода Aedes
Aedes caspius и A. dorsalis. В качестве дополнительных диагностических признаков Беккер с соавторами (Becker et al., 2003): указывают также, что у A. caspius главная шиповидная щетинка сильно изогнута на вершине, т.о. вершина оказывается отогнутой назад почти на половину длины щетинки; у A. dorsalis главная шиповидная щетинка слабо изогнута на вершине, т.о. вершина оказывается отогнутой назад менее чем на треть от длины щетинки. Также Беккер с соавторами (Becker et al., 2003) и Мориг (Mohrig, 1969) отмечают, что базальная бородавка у A. caspius без отчетливого уступа, ее верхний край переходит вверх во внутреннюю поверхностью вальвы. В то же время у A. dorsalis базальная бородавка отчетливо выступающая, верхний край образует выраженный угол с внутренней поверхностью вальвы.
Нами при просмотре коллекционных экземпляров у некоторых особей отмечено наличие 3-й дополнительной щетинки, расположенной дорсальнее главной шиповидной щетинки (рис. 28); расстояние между ними может сильно варьировать. Вероятно, четких диагностических признаков, разделяющих A. caspius и A. dorsalis, к настоящему моменту не обнаружено.
Aedes communis и A. pionips. Некоторые авторы (Carpenter, LaCasse, 1955) указывают, что гофрированная форма крыла класпета характерна только для Aedes communis, а у A. pionips крыло плоское, не гофрированное. Данные исследователи также предлагают и другие признаки: соотношение размеров щупиков и хоботка и строение базальной бородавки коксита. Так, у A. communis щупики значительно длиннее хоботка, у A. pionips – короче или равной длинны. Базальная бородавка A. communis сравнительно большего размера, чем таковая A. pionips, на вершинном крае несет утолщенные щетинки. Беккер с соавтарами (Becker et al., 2003) отмечают, что данные щетинки сильнее изогнуты у A. communis, чем у A. pionips.
Нами исследован лишь 1 экз. A. pionips, у которого крыло класпета оказалось также гофрированным, как и у A. communis. Базальная бородавка у исследованного экземпляра была деформирована из-за усыхания, щетинки на ней изогнуты примерно также, как и у A. communis. Для того, чтобы оценить диагностическую ценность данных признаков более достоверно, вероятно, необходимо исследовать большее число точно определенных экземпляров A. communis и A. pionips.
Aedes cataphylla и A. leucomelas. В качестве дополнительных диагностических признаков Беккер с соавторами (Becker et al., 2003): указывают также, что у A. cataphylla склеротизованный боковой участок крыла класпета заметно расширен в какой-либо части крыла, а у A. leucomelas он более узкий, примерно одинаковой ширины на протяжении всего крыла. Мориг (Mohrig, 1969) также отмечает, что у A. leucomelas шиповидная щетинка базальной бородавки на вершине крючковидно изогнута, а у A. cataphylla – нет. Кроме того, у A. cataphylla щетинки лопастей 9-го тергита расположены по краю лопасти, у A. leucomelas – также и на поверхности лопасти. Гуцевич и др. (1970) указывают на строение щетинок на лопастях 9-го тергита, которые обычно короткие и присутствуют в числе от 6 до 10 у A. cataphylla и относительно длинные в количестве от 10 до 15 у A. leucomelas.
Результаты нашего исследования гениталий самцов A. cataphylla и A. leucomelas фондовых коллекций ЗИН РАН показали, что количество и расположение щетинок на лопастях 9-го тергита изменчиво и вряд ли может быть надежным диагностическим признаком, разделяющим данные виды. Однако, у большинства изученных особей A. cataphylla щетинки действительно были направлены назад, а у A. leucomelas – назад и наружу. Различия между данными видами по строению крыла класпета, указанные Беккер с соавтарами (Becker et al., 2003), нами обнаружены не были. Также не были подтверждены различия в строении шиповидной щетинки базальной бородавки коксита, описанные Моригом (Mohrig, 1969). Тем не менее, согласно результатам нашего исследования, длина крыла класпета у A. cataphylla примерно в 3 раза превышает свою наибольшую ширину, а у A. leucomelas – более чем в 4 раза. Поскольку измерения проводились лишь у небольшого числа особей, полученные данные следует рассматривать как предварительные для диагностики A. cataphylla и A. leucomelas.
Aedes sticticus и A. nigrinus. В качестве дополнительных диагностических признаков Беккер с соавторами (Becker et al., 2003): указывают также, что вершинная бородавка у Aedes sticticus сильнее выступает в дистальной части, а у A. nigrinus она сильнее выступает в срединной части. Мориг (Mohrig, 1969) указывает также, что нижний край вершинной бородавки A. sticticus равномерно переходит во внутреннюю поверхность вальвы. У A. nigrinus нижний край вершинной бородавки четко ограничен от внутренней поверхности вальвы.
Результаты нашего исследования гениталий самцов у данных видов фондовых коллекций ЗИН РАН показали, что форма вершинной бородавки на бальзамном микропрепарате сильно зависит от фиксации материала: положения коксита и давления покровного стекла. В связи с этим использование данного признака для диагностики, на наш взгляд, выглядит проблематичным. Признаки базальной бородавки, описанные Моригом (Mohrig, 1969), нами обнаружены не были.
Aedes excrucians и A. euedes. В качестве признаков, различающих данные виды, Беккер с соавторами (Becker et al., 2003) указывают, что вершинная бородавка у A. excrucians небольшая, в дистальном направлении не доходит до уровня сочленения стиля и коксита. У A. euedes, наоборот, вершинная бородавка заходит назад за место сочленения стиля и коксита.
Результаты нашего исследования гениталий самцов A. excrucians и A. euedes
так же, как и в случае с A. sticticus и A. nigrinus, показали, что форма вершинной бородавки на бальзамном микропрепарате сильно зависит от фиксации материала. Поэтому использовать данный признак для диагностики, на наш взгляд, следует с большой осторожностью.
Обсуждение
Генитальный аппарат самцов рода Aedes характеризуется высокой сложностью строения, вместе с тем признаки структур данного комплекса используются для видовой диагностики. Проведенный нами ранее (Халин, 2006) анализ литературных данных по морфологии гениталий самцов сем. Culicidae показал, что для многих его структур имеются синонимичные названия, также существуют некоторые омонимы. Большое число дублирующих названий усложняет терминологию структур гениталий самцов и мешает работе специалистов. Например, термин «базальная бородавка» использовался для обозначения выступа у основания коксита (рис. 18-32), а также класпетов (парных придатков, сочлененных с 9-м стернитом, рис. 4, 7-17). Для выступа у основания коксита кроме названия «базальная бородавка» различные авторы использовали и другие названия (например, «basal lobe» - «базальная доля, лопасть»), что существенно затрудняет работу с литературой. В настоящей работе базальной бородавкой коксита считается более или менее обособленный выступ или лопасть у основания коксита на его внутренней поверхности. Класпеты же представляют собой парные образования, сочлененные с 9-м стернитом брюшка; у некоторых видов (например, в подродах Ochlerotatus и Finlaya рода Aedes) класпеты разделены на проксимальный отдел (стволик) и дистальный (крыло). Именно такие структуры считаются класпетами (Гуцевич и др., 1970), гомологичные им неразделенные образования с щетинками на вершине (например, в подродах Aedimorphus и Stegomyia рода Aedes) назывались базальными бородавками коксита, что не вполне корректно.
Синонимичные названия отмечены также и для элементов хетотаксии структур гениталий самцов. Так, одни и те же кутикулярные выросты обозначались как «шипы», «шиповидные» или «утолщенные щетинки базальной бородавки коксита» (рис. 22, 23, 25-27). Название «шипы», часто используемое в отечественной литературе (например, Гуцевич и др., 1970), на наш взгляд, кажется не самым удачным. Эти структуры в действительности представляют собой щетинки (рис. 26), отличающиеся от остальных таковых на базальной бородавке большей толщиной. Вместо названия «шип базальной бородавки коксита» нами предлагается употреблять термин «шиповидная щетинка базальной бородавки коксита». Нет принципиальной разницы между шиповидной щетинкой и прочими щетинками базальной бородавки, а по толщине щетинок можно условно выделить несколько категорий. Например, на базальной бородавке коксита Aedes caspius (рис. 25) толщина щетинок, расположенных по краю вентральнее главной шиповидной щетинки, равномерно уменьшается в направлении вентрального края. У данного вида главная шиповидная щетинка не очень четко выделяется на фоне остальных, расположенные рядом с ней щетинки уступают ей по толщине примерно в 2 раза и превосходят по толщине дополнительную шиповидную щетинку.
Применение методов РЭМ для исследования генитального аппарата самцов рода Aedes показало, что его некоторые структуры имеют более сложное строение, чем это представлялось при изучении под СвМ. Основное преимущество РЭМ – это возможность увидеть трехмерную форму объекта без существенных искажений, чего трудно достичь при изучении бальзамных микропрепаратов под СвМ. Структуры сложной формы, как, например, кокситы, класпеты, проктигер, у многих видов рода Aedes деформируются при изготовлении бальзамных препаратов (может сказываться излишняя обработка щелочью, а также давление покровного стекла на объект). Поэтому при исследовании гениталий самцов под СвМ крайне сложно получить корректное представление о реальной конфигурации объекта. В связи с этим описания данных структур, приводимые в литературе, были неполными даже в тех случаях, когда речь шла о структурах, признаки которых имеют важное диагностическое значение. При описании в литературе (например, Штакельберг, 1937; Гуцевич и др., 1970; Becker et al., 2003) гениталий самцов видов подрода Ochlerotatus рода Aedes структуры данного комплекса характеризуются в большинстве случаев как двухмерные образования. В связи с этим и форма, и хетотаксия элементов генитального аппарата оказываются отражены не всегда корректно. Например, коксит рассматривался только под одним ракурсом, снизу, соответственно, неполно характеризовалась и его форма в целом, и форма его выростов. Кроме того, вершинная бородавка часто поворачивается при изготовлении бальзамных микропрепаратов под давлением покровного стекла; нередко деформируется и базальная бородавка коксита.
Наши исследования показали, что коксит у видов подрода Ochlerotatus может иметь различную форму, однако, у большинства изученных нами видов он отдаленно напоминает сильно удлиненный эллипсоид, в большей или меньшей степени изогнутый (рис. 2, 34). Его внутренняя поверхность разделена продольным мембранозным участком на дорсальную и вентральную части. В проксимальной и срединной части коксита данный участок проходит ближе к его вентральной части, у дистального края плавно изгибается дорсально. Таким образом, дистальный край дорсальной части внутренней поверхности коксита формирует лопасть – вершинную бородавку. При этом вершинная бородавка может иметь форму хорошо заметного выступа (например, у A. diantaeus, A. cantans, A. communis и у A. punctor, рис. 2, 34) или только складки (у A. caspius ). Вершинная бородавка весьма различно выглядит под разными углами обзора, что важно учитывать при использовании данного признака в качестве диагностического. У основания коксита кровососущих комаров может располагаться выступ более сложной формы – базальная бородавка (рис. 18-32). Изучение пространственной конфигурации данной структуры у разных видов рода Aedes подрода Ochlerotatus показало, что строение данной структуры у них существенно различается. Так, у некоторых видов она имеет форму более или менее обособленного выступа: в форме трутового гриба (A. caspius, A. dorsalis, A. communis, A. punctor, рис. 25, 27), плоского выступа (A. diantaeus, рис. 3, 22), чашевидная (A. sticticus, рис. 31) или лопасти (A. excrucians, рис. 32). У A. cantans базальная бородавка в виде конусовидного выступа (рис. 19), от которого под углом отходит удлиненная изогнутая лопасть. В виду такой сложности и разнообразия строения базальной бородавки, а также важного диагностического значения признаков данных структур, следует учитывать, что исследование бородавки под одним ракурсом (снизу) дает не всегда полные и корректные результаты. Например, на бальзамных микропрепаратах выглядят сходными по форме базальные бородавки A. caspius и A. behningi, хотя в действительности их строение существенно отличается. Базальная бородавка A. caspius имеет вид четко обособленного выступа в форме трутового гриба, задняя поверхность которой, несущая многочисленные щетинки, ориентирована практически перпендикулярно продольной оси коксита (рис. 27). У A. behningi данная структура устроена несколько иначе, представлена лопастью, немного выступающей дорсально (рис. 24). Дорсальная поверхность, несущая многочисленные щетинки, ориентирована вдоль продольной оси коксита. Таким образом, по строению базальной бородавки коксита A. behningi оказывается ближе к A. flavescens и A. excrucians (рис. 23, 32), чем к A. caspius.
Проведенные нами исследования строения класпетов (Халин, 2008) под РЭМ показали, что пластина крыла класпета, будучи повернутой на больший или меньший угол, выглядит совершенно по-разному. Это связано не только с тем, что крыло ориентировано под различным углом к стволику класпета, но и с неровностью самой пластины крыла (рис. 7-9). В связи с этим имели место различия в описаниях и рисунках крыла класпета, приводимыми авторами для некоторых видов подрода Ochlerotatus (Штакельберг, 1937; Barr, 1958; Mohrig, 1969; Гуцевич и др., 1970; Becker et al., 2003). На крыле класпетамогут присутствовать элементы рельефа, например, у Aedes communis – продольные гребни и борозды (рис. 8, 10). По наличию данной структуры A. communis четко отличается от A. punctor (рис. 14), у которого пластина крыла ровная. Заметные продольные борозды присутствуют на крыле класпета A. annulipes (рис. 17), что, вероятно, можно считать отличительным признаком от A. excrucians. Для уточнения диагностической ценности данного признака необходимо исследование большего числа экземпляров A. annulipes, а также изучение класпетов под РЭМ. Существенно отличаются от крыльев класпетов видов подрода Ochlerotatus таковые структуры Aedes rusticus (подрод Rusticoidus), где гребни на крыле класпета направлены перпендикулярно продольной оси крыла. Также заслуживает внимание такой признак, как равномерность склеротизации крыла класпета. Например, у A. punctor крыло класпета равномерно склеротизовано, однако, у большинства видов в нем имеются более и менее склеротизованные участки. Так, у A. diantaeus слабо склеротизованные участки имеют вид овальных окошечек (рис. 7), которые на бальзамных микропрепаратах выглядят как сквозные отверстия; у большинства же исследованных нами видов крыло класпета представлено продольной сильно склеротизованной частью («стержень крыла класпета») и 1 или 2 латеральными слабо склеротизованными участками («прозрачное расширение крыла», рис. 12, 13, 15).
Интересные данные получены в результате исследования строения лопастей 9-го тергита. В результате изучения внутривидовой изменчивости некоторых видов рода Aedes показано, что число и расположение щетинок на лопастях 9-го тергита варьирует в широких пределах. Например, у A. communis щетинок может быть от 6 до 11, у A. cantans и A. cataphylla – от 6 до 13, у A. leucomelas – от 8 до 14, у A. diantaeus – от 8 до 15. Для некоторых видов интервал, полученный нами, отличался от литературных данных. Например, для A. diantaeus указывается от 6 до 8 щетинок (Becker et al., 2003). Возможно, это связано с тем, что у многих видов утолщенные щетинки располагались в несколько рядов на внешней поверхности и заднем крае лопасти (рис. 36, 37), поэтому посчитать их под СвМ было сложно. Для видовой диагностики, вероятно, большее значение имеет расположение щетинок в несколько рядов, чем их точное количество, которое сильно варьирует. Также изменчивой оказалась и форма самих лопастей, которые выступают дорсально над остальной поверхностью тергита, их форма также неодинакова при рассмотрении под разными ракурсами. Более постоянным оказался признак соотношения длины и толщины щетинок. Например, у A. cantans щетинки намного длиннее, чем у A. diantaeus и A. communis. Также, возможно, сравнительно стабильным признаком можно считать положение лопастей 9-го тергита, то есть расстояние между ними по отношению к ширине лопасти. Данный признак можно использовать как дополнительный для определения близких видов A. cataphylla и A. leucomelas (рис. 36-38) Так, согласно результатам наших исследований, у A. cataphylla лопасти 9-го тергита удалены друг от друга, расстояние между ними примерно равно ширине лопасти. Лопасти 9-го тергита A. leucomelas, наоборот, расположены близко друг к другу, расстояние между ними составляет примерно половину от ширины лопасти.
Структуры проктигера практически не используются для диагностики видов рода Aedes фауны России. Тем не менее, данный комплекс сложно устроен, и его признаки, возможно, имеют некоторую ценность для определения видов кровососущих комаров. Так, нами отмечены интересные особенности проктигера, отличающие A. diantaeus от большинства других представителей подрода Ochlerotatus, характерных для фауны Северо-Запада России. Проктигер A. diantaeus имеет форму рыбьего хвоста, на дорсальной поверхности с обширным мембранозным участком на его дорсальной поверхности, дистальные участки парапроктов изогнуты внутрь и дорсально. Проктигер других видов (A. caspius, A. dorsalis, A. communis, A. cataphylla, A. leucomelas, A. punctor, A. cantans, A. excrucians, A. flavescens, A. behningi, A. cyprius, A. euedes, A. mercurator, A. pullatus и A. sticticus) представлен парными крючковидными сильно склеротизованными образованиями, изогнутые концы которых направлены дорсально.
Важное диагностическое значение, по-видимому, имеют структуры эдеагуса, главным образом строение фаллосомы. У представителей номинативного подрода Aedes, а также подродов Aedimorphus и Stegomyia она разделена на 2 симметричных латеральных участка; в других подродах (например, Ochlerotatus, Finlaya и Rusticoidus) фаллосома цельная (Reinert, 2000). Данный признак был одним из основных, по которым Рейнерт (Reinert, 2000) предложил разделить род Aedes на 2 рода – номинативный и Ochlerotatus. Нами было отмечено, что у некоторых видов подрода Ochlerotatus фаллосома не полностью разделена на латеральные участки, на ее дистальном крае имеется более или менее глубокая выемка (рис. 33). Наиболее глубокая выемка у Aedes diantaeus, она составляет 2/3 длины фаллосомы. Кроме того, при изучении генитального аппарата представителей подрода Rusticoidus оказалось, что фаллосома у них довольно сложно устроена: имеется медиальная удлиненная склеротизованная часть и 2 латеральных участка. Данные признаки не указывались в качестве диагностических, хотя фаллосома видов подрода Rusticoidus существенно отличается от таковой подрода Ochlerotatus.
Заключение
Подробное исследование гениталий самцов 29 видов из 5 подродов рода Aedes, находки которых возможны на территории Северо-Запада России, позволило нам обнаружить некоторые дополнительные диагностические признаки. К таким признакам можно отнести общую трехмерную форму коксита и, в частности, его базальной бородавки. Форма базальной бородавки часто использовалась для диагностики видов рода Aedes (например, для разделения таких близких видов, как A. caspius и A. dorsalis, A. nigrinus и A. sticticus, A. cantans и A. riparius). Однако, как уже было отмечено выше, в данных случаях принималась во внимание лишь двухмерная форма данной структуры снизу, то есть то, как она выглядит на бальзамном микропрепарате. Так как и коксит в целом, и базальная бородавка имеют сложную пространственную организацию, важно рассматривать их под разными ракурсами. По результатам нашего исследования были уточнены признаки данной структуры, разделяющие такие виды, как Aedes rusticus, A. diantaeus, A. intrudens, A. pullatus, A. caspius, A. dorsalis, A. nigripes, A. impiger, A. communis, A. pionips, A. punctor, A. hexodontus, A. cataphylla, A. leucomelas, A. sticticus, A. nigrinus, A. cantans, A. riparius, A. flavescens, A. mercurator, A. cyprius, A. behningi, A. annulipes, A. excrucians и A. euedes.
Признаки класпетов также использовались для диагностики не всегда корректно, поскольку описывались как плоские фигуры. В определительной таблице класпеты характеризовались нами в качестве объемных образований, в результате чего отмечены признаки, разделяющие виды A. diantaeus, A. intrudens и A. pullatus, A. communis, A. punctor и A. hexodontus, A. annulipes, A. excrucians и A. euedes, а также некоторые другие. В частности, нами отмечено, что крыло класпета A. communis существенно отличается от такового у близких видов (A. punctor и A. hexodontus) наличием четко выраженных продольных гребней.
Следует упомянуть о структурах, признаки которых были заявлены как диагностические, но таковыми, возможно, не являются. Например, количество утолщенных щетинок на лопастях 9-го тергита нередко варьирует в широких пределах внутри одного вида. Этот признак указан в качестве диагностического для разделения видов A. cataphylla и A. leucomelas (Гуцевич и др., 1970). Также, на наш взгляд, не следует использовать для диагностики некоторые признаки формы шиповидных щетинок базальной бородавки коксита [например, у A. caspius и A. dorsalis, отмеченные Беккером и соавторами (Becker et al., 2003), а также у A. cataphylla и A. leucomelas (Mohrig, 1969)]. Кроме того, нами не было замечено различие в строении крючковидно изогнутых щетинок, которые согласно Беккеру и др. (Becker et al., 2003) у A. communis изогнуты сильнее, чем у A. pionips. Для того чтобы оценить диагностическую ценность этих признаков более достоверно, необходимо исследовать генитальный аппарат у большего числа точно определенных экземпляров A. communis, A. pionips, A. caspius, A. dorsalis, A. cataphylla и A. leucomelas. Так как нет надежных признаков для различения этих видов ни для какой фазы жизненного цикла, то получить достоверно определенный материал оказывается затруднительным. Нет надежных диагностических признаков для разделения и ряда других видов (например, A. punctor и A. hexodontus, A. excrucians и A. euedes).
Таким образом, приведенный в настоящей работе вариант определительной таблицы следует рассматривать как предварительный, требующий дальнейшей работы. Некоторые виды исследовались нами лишь в небольшом числе экземпляров, вследствие чего точно охарактеризовать пределы внутривидовой изменчивости признаков генитального аппарата не удалось.
Литература
Гуцевич А.В., А.М. Дубицкий Новые виды комаров фауны Советского Союза. Паразитол. сб. 1981. Т. 30. С. 97-165.
Гуцевич А.В., Мончадский А.С., Штакельберг А.А. Комары, семейство Culicidae. Фауна СССР. Насекомые двукрылые. Л., 1970. Т. 3, вып.4. 384 c.
Халин А.В. Диагностические признаки коксита в гениталиях самцов комаров Aedes caspius (Pallas), A. cantans Meigen и A. communis (De Geer) (Diptera, Culicidae) // Энтомол. обозр. 2005. Т. 84, вып. 4. С. 884-896.
Халин А.В. Строение генитального аппарата самцов Aedes diantaeus Howard, Dyar et Knab (Diptera, Culicidae)) // Энтомол. обозр. 2006а. Т. 85, вып. 4. С. 737-748.
Халин А.В. Диагностические признаки комаров рода Aedes фауны России (Diptera, Culicidae) // Автореферат диссертации на соискание ученой степени кандидата биологических наук. Санкт-Петербург, 2007. 24 с.
Халин А.В. Строение класпетов в гениталиях самцов комаров подрода Ochlerotatus рода Aedes и их диагностические признаки (Diptera, Culicidae) // Энтомол. обозр. 2008…
Халин А.В., Горностаева Р.М. К таксономическому составу кровососущих комаров (Diptera: Culicidae) мировой фауны и фауны России (Критический обзор) // Паразитология. 2008. Т. 42, вып. 5. С. 360-381.
Штакельберг А.А. Кровососущие комары Палеарктики. М.; Л., 1937. 257 c.
Barr A.R. The mosquitoes of Minnesota (Diptera: Culicidae: Culicinae) //Technical Bulletin. 1958. Vol. 228. 154 p.
Becker N., Petric D., Zgomba M., Boase C., Dahl C., Lane J., Kaiser A. Mosquitoes and their control. Kluwer Academic / New York, Boston, Dordrecht, London, Moscow: Plenum Publishers, 2003. 498 p.
Belkin J.N. The mosquitoes of the South Pacific (Diptera, Culicidae). Berkeley and Los Angeles: University of California Press, 1962. Vol. 1. 608 p.
Belkin J.N. The mosquitoes of the South Pacific (Diptera, Culicidae). Berkeley and Los Angeles: University of California Press, 1962. Vol. 2. 412 p.
Carpenter S.J., La Casse W.J. Mosquitoes of North America (north of Mexico). University of California Press, 1955. 360 p.
Edwards F.W. Diptera, fam. Culicidae. Genera Insectorum. Brussels, 1932. Fasc. 194. 258 p.
Knight K.L., Laffoon J.L. A mosquito taxonomic glossary. V. Abdomen (Except female genitalia) // Mosquito Systematic Newsletter. 1971. Vol. 3. N 1. P. 8-24.
Mohrig W. Die Culiciden Deutschlands // Parasitolog. Schriftenreihe. 1969. H. 18. 100 s.
Reinert J. F. Restoration of Ayurakitia to generic rank in tribe Aedini and a revised definition of the genus // Journal of the American Mosquito Control Association. 2000a. V. 16, N 2. S. 57-65.
Reinert J. F. New classification for the composite genus Aedes (Diptera: Culicidae: Aedini), elevation of the Ochlerotatus to generic rank, reclassification of the other subgenera, and notes on certain subgenera and species // J. Amer. Mosquito Control Assoc. 2000. Vol. 16, N 3. P. 175-188.
Reinert J. F., Harbach R. E., Kitching I. J. Phylogeny and classification of Aedini (Diptera: Culicidae), based on morphological characters of all life stages // Zoological Journal of the Linnean Society. 2004. V. 142. S. 289-368.
Reinert J. F., Harbach R. E., Kitching I. J. Phylogeny and classification of Finlaya and allied taxa (Diptera: Culicidae: Aedini) based on morphological characters of all life stages // Zoological Journal of the Linnean Society. 2006. V. 148. S. 1-101.
Reinert J. F., Harbach R. E., Kitching I. J. Phylogeny and classification of Finlaya and allied taxa (Diptera: Culicidae: Aedini) based on morphological characters of all life stages // Zoological Journal of the Linnean Society. 2006. V. 148. S. 1-101.
Reinert J.F., Harbach R.E., Kitching I.J. Phylogeny and classification of Ochlerotatus and allied taxa (Diptera: Culicidae: Aedini) based on morphological data from all life stages // Zoological Journal of the Linnean Society. 2008. V. 153. S. 29–114.
А.В. Халин
Январь 2009 г.





.JPG)































