Общая характеристка трематод (Trematoda, или Digenea) |
||
АвторыРедактор |
Трематоды (Trematoda, или Digenea) – подкласс плоских червей, ведущих облигатно эндопаразитический образ жизни. Насчитывая более 2700 номинальных родов и около 18000 видов, трематоды являются одной из наиболее многочисленных и широко распространенных групп паразитических беспозвоночных. Для трематод характерен сложный жизненный цикл, протекающий со сменой двух или трех хозяев – первого промежуточного, второго промежуточного (дополнительного) и окончательного. Первыми промежуточными хозяевами являются брюхоногие или двустворчатые моллюски, в редких случаях ими могут быть полихеты. В роли второго промежуточного хозяина могут выступать самые разные животные (рыбы, земноводные, моллюски, олигохеты, пиявки, членистоногие и др.). Окончательными хозяевами всегда являются позвоночные. В теле позвоночных трематоды, как правило, локализуются в пищеварительном тракте и связанных с ним железах, однако могут населять и другие органы и ткани, включая легкие, почки, кровеносную систему, полость тела и половые протоки. Около 12 видов трематод паразитирует в человеке; самыми распространенными и опасными из таких паразитов являются шистосомы. Вызываемое шистосомами заболевание (шистосоматоз) встречается в 74 странах мира и уносит ежегодно около полумиллиона человеческих жизней. |
Содержание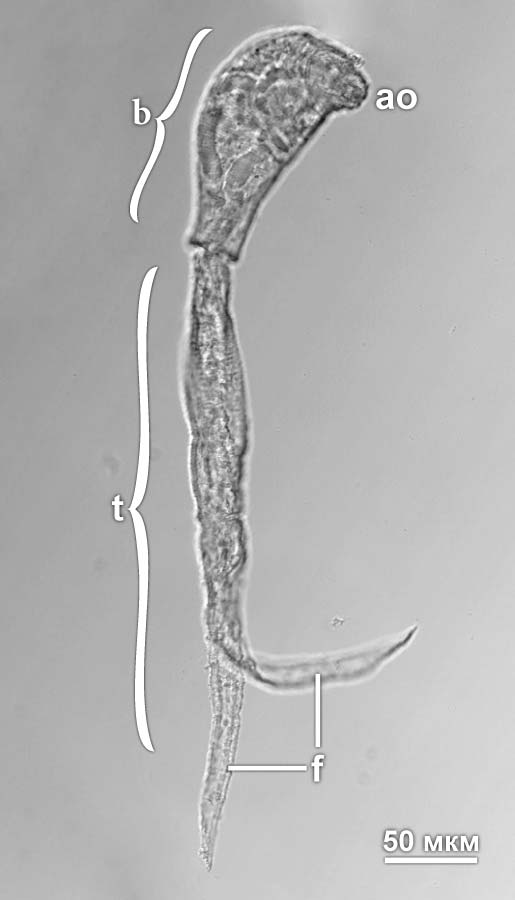
Рис. 1. Церкария Diplostomum pseudospathaceum. Обозначения: ao – передний орган (железисто-мышечный орган, превращающийся у метацеркарий в ротовую присоску); b – тело церкарии; f – фурка хвоста; t – хвост. Фото А.А. Петрова

Рис. 2. Обобщенная схема организации мариты. Обозначения: ac – брюшная присоска (ацетабулюм); at – половой атриум; cr – циррус; cs – мешок цирруса; ec – выводящий канал; eo – выделительное отверстие; ex – мочевой пузырь; gb – ветвь кишечника; mg – железы Мелиса; mo – ротовое отверстие; ph – глотка; pp – предглотка; os – ротовая присоска; ot – оотип; ov – яичник; sr – семяприемник; te – семенник; vc – желточный проток; vf – желточный фолликул.
|
Чередование поколений |
Развитие трематод протекает с чередованием поколений. Жизненный цикл обычно включает два партеногенетических и одно гермафродитное поколение. Отложенные половозрелыми трематодами (маритами) яйца выводятся во внешнюю среду, где из них выходят свободноплавающие ресничные личинки первого партеногенетического поколения – мирацидии. Мирацидий проникает в первого промежуточного хозяина, сбрасывая при этом ресничные клетки, и претерпевает метаморфоз, превращаясь в партеногенетическую особь – материнскую спороцисту. Спороциста имеет довольно простое строение и представляет собой образованный стенками тела мешок. В центральной полости мешка развиваются эмбрионы, которые дают начало редиям, особям второго партеногенетического поколения. Редии устроены сложнее, чем спороцисты, имеют функционирующую пищеварительную систему и могут активно передвигаться в теле хозяина. У некоторых трематод поколение редий отсутствует, и материнские спороцисты дают второе поколение спороцист, так называемые дочерние спороцисты. Развивающиеся в редиях или дочерних спороцистах эмбрионы превращаются в церкарий, которые покидают тело промежуточного хозяина. Церкария (рис. 1), являющаяся личинкой гермафродитного поколения трематод, несет на заднем конце мускулистый хвост, при помощи которого она передвигается в толще воды. У большинства трематод церкарии самостоятельно находят второго промежуточного хозяина, проникают в него, отбрасывая при этом хвост, и превращаются в метацеркарий. Второй промежуточный хозяин вместе с метацеркариями проглатывается окончательным хозяином, в нем метацеркарии развиваются в мариты. В некоторых группах трематод второй промежуточный хозяин отсутствует и церкарии проникают непосредственно в окончательного хозяина. У представителей примитивных семейств личинки инцистируются на водной растительности и остаются в состоянии покоя, пока их не проглатывает окончательный хозяин. |
|
Строение мариты |
Тело мариты (рис. 2) обычно имеет уплощенную, листовидную форму. Размеры большинства марит не превышают 5 мм, но среди трематод встречаются и настоящие гиганты, такие как Hirudinella ventricosa, вырастающая до 12 см в длину. Исключительных размеров достигают нитевидные мариты из семейства Didymozoidae – паразитирующая в луне-рыбе дидимозоида Nematobibothrioides histoidii может вырастать в длину до 12 метров. Марита обычно несет две присоски – ротовую, которая окружает ротовую полость, и брюшную (ацетабулюм), но у некоторых трематод одна или обе присоски могут отсутствовать. Тело мариты покрыто синцитиальным тегументом, поверхность которого, как правило, вооружена шипиками, состоящими из кристаллизованного актина. Пространство между внутренними органами заполнено паренхимой, в которой у некоторых трематод образуется сеть каналов, называемая лимфатической системой; эта система по всей видимости выполняет транспортную функцию. Мышцы стенки тела, как правило, представлены тремя слоями – наружным кольцевым, промежуточным продольным и внутренним диагональным, однако встречаются различные вариации в организации мускулатуры. Пищеварительный тракт слепо замкнут и обычно состоит из короткой предглотки, мышечной глотки, пищевода и двух (в редких случаях одной) ветвей кишечника. У некоторых трематод ветви кишечника замыкаются в кольцо. Выделительная система представлена протонефридиями; протонефридии собираются в общие выводные каналы, которые ведут в мочевой пузырь, открывающийся наружу выделительным отверстием в задней части тела. У крупных форм выстилка мочевого пузыря может образовывать в паренхиме сеть сосудов (паранефридиальный плексус), которая выполняет метаболическую или экскреторную функцию. Половая система обычно занимает большую часть тела мариты и имеет исключительно сложное строение. Подавляющее большинство трематод гермафродиты, раздельнополость характерна только для шистосоматид и некоторых дидимозоид. Мужская часть половой системы состоит из одного или чаще двух семенников, от которых отходят выводящие протоки, заканчивающиеся копулятивным органом – циррусом. Женская половая система включает яичник, желточники (желточные фолликулы) и сложную систему каналов и желез, участвующих в образовании оболочек яйца. Выходящий из яичника зрелый ооцит попадает в расширенную часть яйцевода – оотип, в котором происходит оплодотворение и начинается формирование яйца. Оотип окружен железами Мелиса, секрет которых служит матрицей для формирующихся скорлуповых оболочек и вероятно также образует смазку, облегчающую продвижение яйца по половым протокам. В оотип также открывается желточный проток, по которому к формирующемуся яйцу поступают желточные клетки. Желточные клетки образуют скорлуповую оболочку и обеспечивают питание для развивающегося эмбриона. Дальнейшее формирование яйца происходит в матке, которая открывается наружу в передней части тела, причем ее отверстие часто располагается вместе с мужским половым отверстием на дне общего полового атриума. |
|
Органы чувств |
Наиболее характерными органами чувств трематод, присутствующими на всех стадиях их жизненного цикла, являются многочисленные рецепторные образования в тегументе. Они крайне разнообразны по своему строению и могут быть представлены одноресничными сенсиллами, ресничными ямками или нересничными чувствительными папиллами. Их рецепторная функция, по всей видимости, различна. Среди них могут быть хемо-, механо-, термо- или реорецепторы, однако углубленные исследования их функционального назначения у трематод до настоящего времени не проводились. Рецепторные образования тегумента обычно рассеяны по всей поверхности тела, но наиболее густо расположены на присосках и органах проникновения. Их расположение часто видоспецифично и активно используется в диагностике видов. У большинства мирацидиев и у многих церкарий имеется по два или три инвертированных фоторецепторных глазка. Глазки состоят из рабдомерных рецепторных и меланин-содержащих пигментных клеток. Для некоторых мирацидиев и церкарий характерны пластинчатые ресничные органы, которым также приписывается фоторецепторная функция. |

Рис. 3. Серотонинергическая нервная система метацеркарии Diplostomum pseudospathaceum. Окраска антителами к серотонину. Обозначения: an – нервы, иннервирующие брюшную присоску; cg – церебральный ганглий; de – дорсальный нервный ствол; on – нервы, иннервирующие ротовую присоску; tc – поперечные комиссуры; ve – вентральный нервный ствол. Фото А.А. Петрова

Рис. 4. RFамид-ергическая нервная система метацеркарии Diplostomum pseudospathaceum. Окраска антителами к FMRFамиду. Обозначения: cg – церебральный ганглий; de – дорсальный нервный ствол; le – боковой нервный ствол; on – нервы, иннервирующие ротовую присоску; tc – поперечные комиссуры; ve – вентральный нервный ствол. Фото А.А. Петрова
|
Нервная система |
Нервная система марит имеет форму ортогона, характерного для плоских червей. Мозг расположен над глоткой и состоит из пары церебральных ганглиев, соединенных между собой надглоточной комиссурой. У большинства исследованных видов имеется четыре пары передних нервов; три из них (дорсальная, латеральная и вентральная) отходят от церебральных ганглиев и иннервируют в основном ротовую присоску и сенсорные органы переднего конца тела, четвертая пара начинается от комиссуры мозга и иннервирует глотку. Назад от церебральных ганглиев отходит шесть стволов, образующих вентральную, латеральную и дорсальную пары. Вентральные продольные стволы обычно развиты сильнее двух других пар, идут вдоль всего тела мариты и у многих трематод соединяются петлей на заднем конце. Латеральные и дорсальные стволы часто короче вентральных и в некоторых группах трематод могут частично или полностью редуцироваться. Продольные нервные стволы соединены между собой расположенными через регулярные промежутки поперечными комиссурами. Стволы и комиссуры соединены с комплексом периферических плексусов, иннервирующих мускулатуру стенки тела и внутренние органы. В некоторых группах трематод тело мариты разделяется на два отдела – переднее и заднее тела; последнее содержит органы половой системы. У таких марит в задней части переднего тела имеется дополнительный ганглий, иннервирующий заднее тело. Строение нервной системы церкарий и метацеркарий (рис. 3 и рис. 4) в целом такое же, как у марит. Хвост церкарий обычно имеет хорошо дифференцированную нервную систему: у одних церкарий продольные нервные стволы переходят непосредственно в хвост, у других они заканчиваются плексусом в задней части тела, а от этого плексуса назад отходят две пары хвостовых нервов. Нервная система мирацидия включает крупный, часто двухлопастной, церебральный ганглий и пару нервных стволов. У спороцист нервная система развита слабо, но у некоторых трематод в спороцистах описан церебральный ганглий и одна пара продольных нервных стволов. Редии имеют более сложную нервную систему, чем спороцисты; кроме церебрального ганглия, у них также могут быть развиты две пары передних нервов и две пары продольных нервных стволов. Для нервной системы трематод характерны в основном биполярные и мультиполярные клетки. Основным нейромедиатором, оказывающем возбуждающее действие на их мускулатуру, по имеющимся данным, является серотонин (рис. 3), а основным тормозящим нейромедиатором – ацетилхолин. Это отличает паразитических червей от большинства других изученных к настоящему времени беспозвоночных и позвоночных животных, у которых ацетилхолин оказывает возбуждающее действие на мышечные клетки. В нервной системе трематод обнаружены также нейропептид F и FMRFамид-подобные пептиды (рис. 4). Предполагается, что они принимают участие в регуляции мышечных сокращений. |
|
Рекомендуемая литература |
|
|